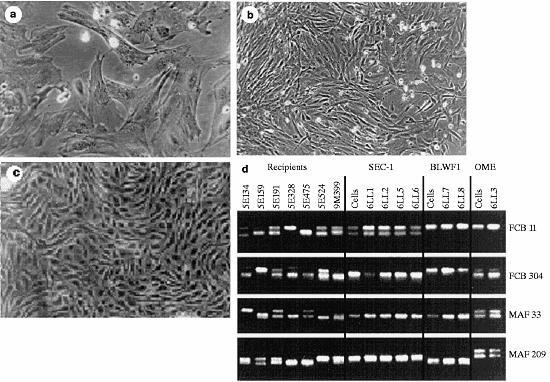
| Tipo celular | No de pares fundidos (%)* | No de recuperados de oviduto (%) | No cultivado | No de mórulas/ blastocistos (%) | No de mórulas ou blastocistos transferidos | No de gestações/ No de receptoras (%) | No de cordeiros vivos (%) |
| Epitélio mamário | |||||||
| Fibroblasto fetal | |||||||
| Derivada de embrião |

| Tipo celular | Raça do cordeiro | Identidade do cordeiro | Duração da gestação (dias)* | Peso ao nascer (kg) |
| Epitélio mamário | Finn Dorset | |||
| Fibroblasto fetal | Black Welsh
Black Welsh Black Welsh |
|||
| Derivada de embrião | Poll Dorset
Poll Dorset Poll Dorset Poll Dorset |
| Texto traduzido por:
João M. P. Alves (Jota) em 17/3/97 e-mail: [email protected] Visite minhas páginas na Web! http://www.geocities.com/CapeCanaveral/Lab/3891/ (acho que você acabou de passar por ela...) http://www.geocities.com/SunsetStrip/Towers/6956/ (é a página do Rising Sun, minha banda de rock, com MIDIs, links e letras das melhores bandas de rock do planeta). |