it may take quite a while to complete it!!

| ALL _THE _INFORMATION _YOU _WANT _TO _KNOW _______ ON_ THE_ FOSSA_ CRYPTOPROCTA_ FEROX________ |
Cryptoprocta bennett 1833:46. Type species Cryptoprocta ferox Bennett, 1833, by original designation.
Viverridae, Subfamily Cryptoproctinae. The subfamily includes a single living species.
Fossa Cryptoprocta ferox Bennett, 1833:46. Type locality "from madagascar." Cryptoprocta typicus A. Smith, 1834:34 Type locality
Madagascar.
Context noted in genetic summary above. No subspecies have been recognized.
Cryptoprocta differs externally from other canivores of Madagascar in being considerably larger and cat-like (fig. 1). The combined
character of uniformly colored pelage, large size (largest of viverridae other than macrogalidia and viverra), long tail (slightly shorter
than head and body), semiplantigrade feet, retractile claws, rounded external ear, and relatively short face render it distinctive among
viverrids. C. ferox somewhat resembles a small, short-legged, dark-colored puma (felis concolor).
Published measurements are few, and for most the method of measurement and the source of information are omitted, Measurements
(in mm "millimeter") of adult specimens are: length of head and body, 700 to 800; tail 650 to 700; hindfoot, 120 to 128; body mass
ranges from 7 to 12 kg. Other measurements (in mm) of head and body are 600 to 750 and 850 to 900. listed the height at the shoulder
as 370mm. Our measurements of mounted specimens in Rijksmuseum van Natuurlijke Historie, Leiden, indicated that the external ear
was about 45mm long and the hindfoot about 142mm. Body size increase from the east to the southwest in Madagascar. Males are
slightly larger than females. The reddish-brown pelage blends to grayish on the dorsum; the underparts are nearly beige, and the tail
is paler than the flanks. The pelage is without pattern or spots and it resembles the winter pelage of the European roe deer
(Capreolus capreolus), but with a reddish tinge. Published details of the coloring of C. ferox differ greatly; however, all authors
acknowledge the predominance of reddish brown. stated that the underparts of males in rut are stained reddish, similar to that on
male red kangaroos (Macropus rufus), although Albignac (1973) was unable to comfirm this observation. Darker fossas, probably are
melanistic rather than subspecifically distinct as proposed by Kaudern (1915). The meonatal pelage is gray or nearly white, but adult
coloration is attained after 6 months. Body hairs are relatively short, straight, and dense.
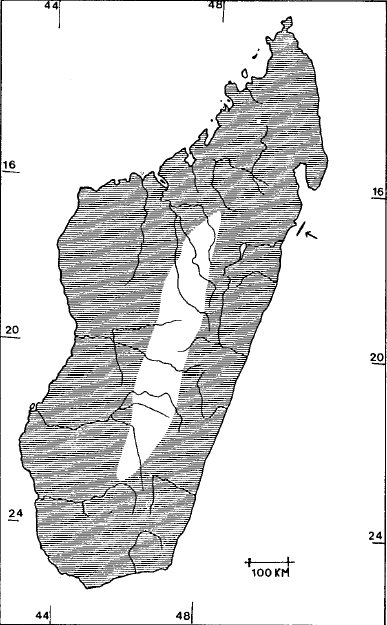
Cryptoprocta ferox occurs throughout Madagascar and on the small island of Saint-Marie (fig. 2) off the east coast of Madagascar.
On Madagascar, fossas are common near Bongolava (in the NW near Majunga), on the E coast near Morondava, and on the NE
coast near Antalaha. They occur in forested areas including mountain to 2,000m (Adringitra Mountains), but avoid treeless habitats
(central highlands).
Grandidier (1902) descrided material from the cave of Andrahomana, 40 km S of Fort Dauphin in southern Madagascar as a new
subspecies, C. F. spelea. petit (1935) redescribed the same subfossil cave material, but regarded it as a separate species and applied
the name C. spelea to it. He described it on the premise that the earlier appellation was a nomen mudum. We consider the earlier name
valid because Grandidier (1902:592) gave a short but clear description "de grande taille," and thereby followed the international Code
of Zoological Nomenclature (1958: Art. 12, 32[a], 45[e]i). Lamberton (1939) described the specie, Cryptoprocta antamba, from subfossil
material from near Triandroina in southern Madagascar. Lamberton (1939) joined petit (1935) in giving specific rank labelled C. spelea
on deposit in the Museum National d'Historie Naturelle, paris (nos. 1977-755 and 1977-756), have condylobasal lengths of 152 and
138mm, respectively. In our opinion, the cranial differences betweeb C. ferox and C. spelea, as understood and described by
Lamberton (1939), can be explained by allometric changes in C. spelea with growth; therefore, the validity of C. spelea as a separate
species is questionable. Skulls, teeth, and postcranial materials identified as C. ferox, C. spelea, and C. antamba from different caves in
central, southern and southwestern Madagascar give no indication of the phylogeny of the Recent population because they are
inseparable morphologically. Palaeoprionodon Filhol, 1880 (upper Eocene or lower Oligocene-Miocene, Europe) also has many
viverrid-like characters but is a member of the Felidae Conversly called C. ferox a viverrid with many cat-like characters.
"Cat and viverrids were closely similar in the early Oligocene. . ."
The rhinarium is large and prominent as in other viverrids, but the large and rounded external ear with a lateral bursa resembles that of
Felidae. The conspiciously long facial vibrissae, some even longer than the head (bennett, 1835), are arranged as in other canivores,
but the interramal group in C. ferox and other viverrids is lacking in the Felidae. Carpal vibrissae are present as in some other viverrid
genera. The soles of the feet bear strong pads and are nearly naked. The claws are retractile but not hidden in skin sheaths as in
Felidae. Perianal skin glands are contained in an anal sac surrounding the anus like a pocket as in Herpestinae. The anus lies within
this pocket which opens to the exterior as a horizontal slit below the tail. A sternal gland that supposedly produce a reddish
substance during rut in males. could not be found by Albignac (1973). Other seent glands lie near the penis or vulva and on the neck.
Albignac (1973) mentioned that a strong odor emanates from scent glands near the penis. Prescrotal glands are absent as in
Herpestinae. Scent glands are less devoloped in females. There are three pair of teats; the composition od mikl is unknown. The skull
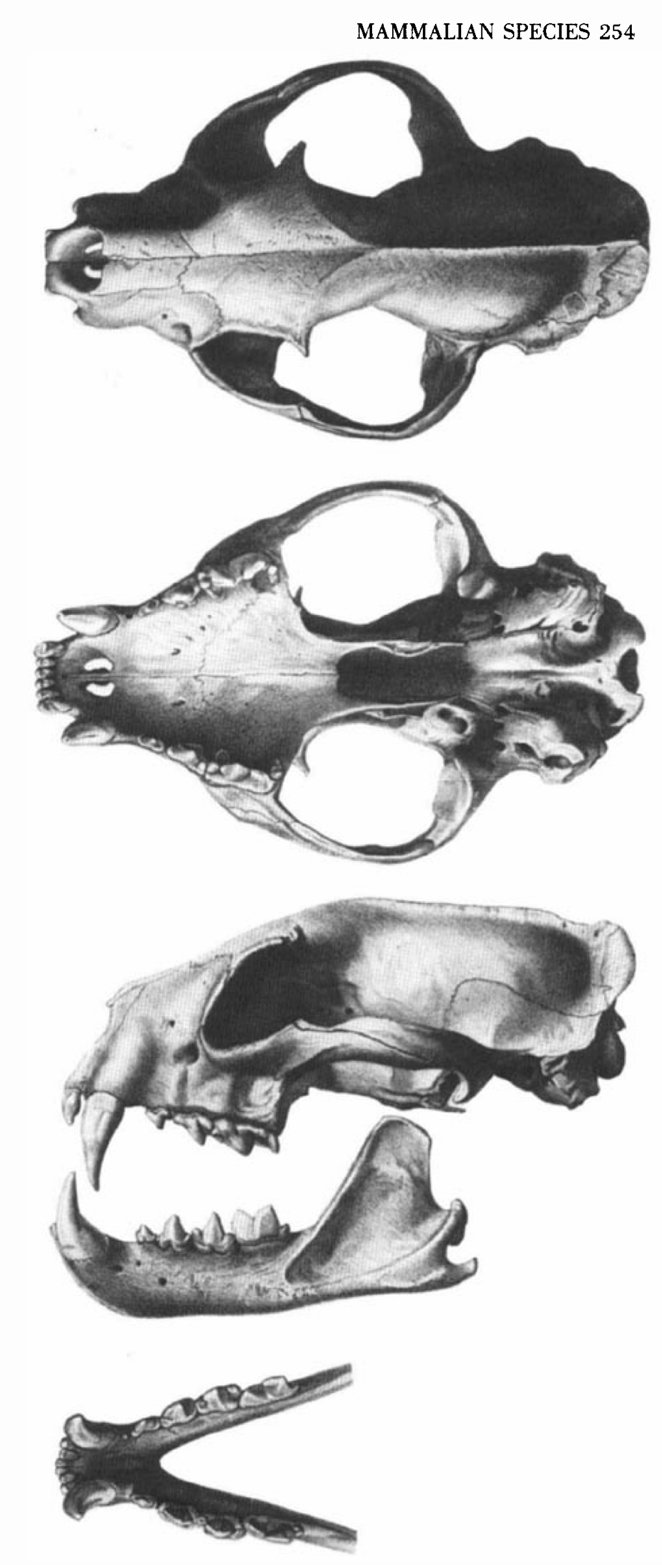
of C. ferox is constricted behind the orbits (Fig. 3 "see it below). The postorbital processes of frontal and jugal are short and do not
meet to separate the orbit and temporal fossa. The alisphenoid extends some distance between the squamosal and frontal bones, a
feature associated with the relatively long braincase and common in other Viverridae. The facial skull is shorter than in almost all other
viverrids, but longer than in any felid. The heavily built masticatory muscles are large and robust at their origins. The sagittal and
lambdoidal crests are well developed. the zigomatic arch extends broadly from the skull, and the internal ptergoid process in long and
supports the action of the strong internal pterygoid muscle. An alisphenoid canal is always present (present in viverridae, except
Galidiinae and sometimes Eupleures and Cynogale, lacking in Felidae and Hyenidae, except sometimes present in Crocuta;. The carotid canal lies entirely within the basisphenoid, but has no contact with the petrosum (as in all Viverridae but none of the Felidae;.
The hypoglossal condyloid foramen is separated from the jugular foramen ("posterior lacerate foramen) by a bony wall as in Galidia
and Felis; in Herpestes, Viverra, and Paradoxurus both foramina form a single opening. The bulla is typical of that of viverrids,
probably exhibiting greatest affinity with that of viverra. The bulla is united (synostosis) with the basisphenoid (fig. 3) as in the
Viverridae but not among Felidae. The tympanic bone (" ectotympanic) lies rostrolaterally and constitutes the smaller part of the bulla;
it is in synostosis with the entotympanic, but is easily distinguished as a separate bone. The tympanic is not inflated and there is no
depression or hole in its ventral surface. The external auditory meatus is large and its rim is more or less smooth and not extended like
a spout (fig. 3). Thus the timpanic is different from that in Herpestinae. The large caudomedial entotympanic forming the larger part of
the bulla is bony, mainly inflated caudally, and flattened on both sides. The paroccipital process resembles that of Herpestinae; it is
flatty attached to the bulla caudally by its whole anterior surface and does not extend ventrally to the bulla. The lower jaw is robust;
its articular process extends farther posteriorly than its coronoid process (fig. 3). The condylobasal length (in mm) for five males
(mean and range) and one female are: 128.7 (115.1 to 139.5) and 117.2, respectively. The dental formula for the permanent dentition is i
3/3, c 1/1, p 3-4/3-4, m 1/1, total 32 to 36, and for the deciduous dentition is i 3/3, c 1/1, p3/3, total 28. Carnissials (especially ml) and
molars have an almost cat-like "grade of advancement" (Gregory and Hellman, 1939), whereas incisors and the remaining premolars are
typical of the viverridae. The upper canine is developed as in Felis, and Pl and pl are small or missing in C. ferox; they are present in
most Viverridae, but not in Felidae. Most authorities refer to these teeth as being of the second dentition, but the first premolars are
not replaced in fissipeds. Both P2 and p2 have two roots (these teeth are missing in felids) and P3 has a talon, as in some other
Viverridae but not in Felidae. The upper carnassial (P4) is large, sectorial, and of nearly the same shape as that in Felidae; M1 is small
(as in Felis) and M2 is missing (as in Felidae, but in contrast to its presence in most Viverridae). The similarity of the lower carnassial
(m1) with that of the Felidae is related to the absence of a matacomid and the greatly reduced talonid; m1 in other Viverridae has a
metaconid and larger talonid. Deciduous incisors are similar to those other Viverridae; the deciduous canine is similar to that in
Herpestes and viverra and unlike that of the Felidae, and dp4 is similar to that in Genetta. The postcranial skeleton presents no
unusual features. The vertebral formula is 7C, 13T. 7L, 3S, and 29 or 30 Ca, total 59 or 60. The clavicle is small, 8mm long in an adult
female, and its fixed by ligaments to the sternum and scapula. The entepicondylar foramen of the humerus is present as in most
non-Malagasy viverrids. The supraspinous fossa of the scapula is larger than the infraspinous one (ratio 1.5:1), as in Galidia
Herpestes, and Felis. The radius is shorter than the humerus and the tibia shorter than the femur. Certain myological characteristics
are of interest for comparative studies: overall strong masticating musculature; existence of an intermediate tendom within the
digastric muscle; long origin of the latissimus dorsi over 14 vertebrae; existence of teres minor and supinator brevis muscles; and
unsual muscles on the soles of the feet. According to Carlsson (1911), herpestine feature of the brain of C. ferox include the presence
of a crucial sulcus, a short sylvian sulcus (missing in some herpestine species;, and the absence of a presylvian sulcus. Unlike
Herpestinae, there is a suprasylvian sulcus complete posterioly and a postlateral sulcus medially (Radinsky, 1975). Beddard (1895)
reluctantly described a short "fissure" (= sulcus) as the postsylvian, but Carlsson (1910) and Mivart (1885) emphasized that such was
the sylvian sulcus. This agrees with the figures of the brain provided by Brauer and Schober (1970). As typical in carnivores, the left
lung has three lobes and the right lung four. Features of the circulatory system described to date are typical among the feloidea. The
ramification of the aorta also follows the carnivore pattern. The tongue is broad and rounded; Carlsson, (1911) reported two
circumvallate papillae on each side, whereas Beddard (1895) reported three. The paucity of spiny papillae on the tongue and palatal
ridges (eight) are reminiscent of Felidae. C. ferox has no lyssa, in contrast to viverrids and the domestic cat (Felis catus). The two
parts of the thyroid gland are linked by a small isthmus. The stomach resemble that of the domestic cat, being somewhat elongate with
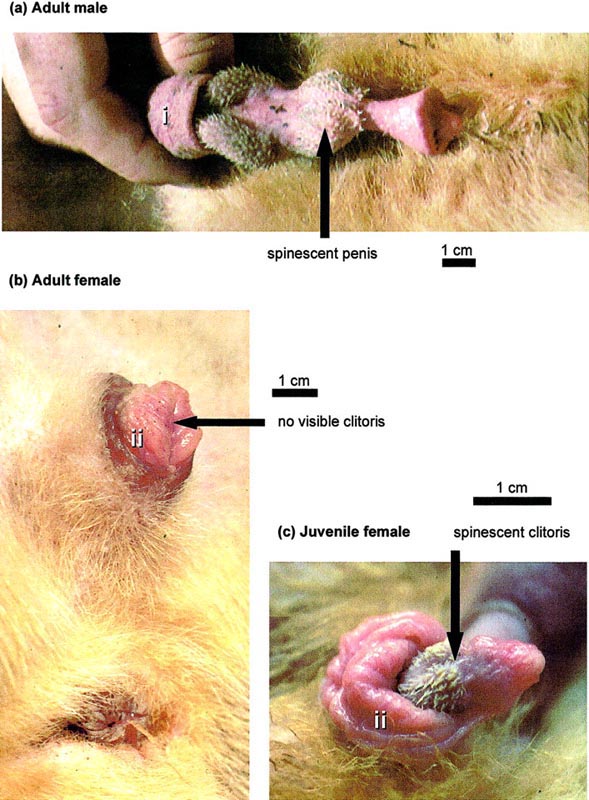
the pyloric part distinctly bent rostrally. The unique structure of the external genitalia in both sexes was the primary reason that
Pocock (1916b) and Lõnnberg (1911) proposed a separate family for C. ferox. Penis and bacalum, are extremely long; erected, the penis
extends forward between the forefeet, and is nearly 20mm thick. The baculum in an adult males is between 58 and 71mm long and is
about 3mm thick. The glans occupies nearly half of the length of the penis and is spiny except at the tip. In felids the glans is
extremely short and spiny in viverrids it is smooth and long. The urethra opens immediately below the tip of the glans. The clitoris is
penis-like, with a well developed preputium and an os clitoridis (about 16mm long and 3mm thick Lõnnberg, 1902) and a pair of
glandular sacs (glands of Bartholini; Carlsson, 1911) that form a scrotum-like structure. As in all Malagasy carnivores, the uterus of
C. ferox is paired throughout its length (duplex uterus); each open into the vagina with a separate cervix. All features of the genitalia
occur in other carnivores, but the mosaic is unique to C. ferox. Structural affinities to other groups of carnivores are as follows: those
features associated with food and its acquisition (facial skull, teeth, tongue, and digestive tract) resemble those of the felids most
closely; the skull, exclusive of the dentition, exhibits greatest resemblance to viverra; some characteristic of the body are most similar
to those of the Herpestinae.
Fossas has very strange penis and clitoris See Picture, originally from this part of www.biolreprod.org ,Also contain more info on the fossa.
{kind=link}
{kind=link}
Fig. 3. Dorsal, ventral , and leteral views of the cranium, and lateral and dorsal views of the mandible of an adult Cryptoprocta ferox
(total length of the skull is 140mm; from Milne-Edwards and Grandidier, 1867).
Fig. 1. Drawing of the Cryptoprocta ferox (from Grassé , 1955) Fossa drawing picture
{kind=link}
Fig. 2. Distribution of Cryproprocta ferox on Madagascar (after Albignac, 1974). The arrow identifies the island of Saint-Marie.
Island picture
{kind=link}
Copulation occurs in September or October in wild C. ferox and parturition occurs in December or January (Albignac, 1969) indicating
a gestation period of about 90 days. Kõhncke (1985) found greater morphological affinities to other viverrids than to felids in the
chondrocrania of fetuses 6 to 7 weeks of age. Birth of tow to four young takes place in a concealed location such as a den or beneath
a tree. Each neonate weight about 100g (or less) and is furred, but toothless and blind (Albignac, 1969). Contrary to a statement by
Albignac (1973), siblings are not always of the same sex. Observations in zoos reveal that postnatal development is slow. Eyes open
after 2 to 3 weeks, the young first leave the den after 4.5 months, then weaning take place. Permanent dentition appears before 20
months of age. Onset of sexual maturity is not before 3. There is a longevity record of 17 years from "Zoologischer Garten" in
Hamburg, Germany, and a male of over 20 years is still living in the zoo at Duisburg, Federal Republic of Germany.
Cryproprocta ferox is generally regarded as the main predator of lemurs, but the evidence is based on a single field observation by
woodcutters. Many foods are taken in captivity: insects, amphibians, reptiles, birds, and small to medium-sized mammals, including a
ring-tailed lemur (Lemur catta; Albignac, 1973). In some accounts. C. ferox is described as a fierce predator on domestic animals,
especially poultry, but also on goats and small calves. In the wild C. ferox probably preys on other viverrids, but it seems to have no
predators of its own other than man. Cryptoprocta ferox lives in virtually all climates on Madagascar and has no special habitat
requirements axcept the presence of trees; it avoids open treeless regions. Unlike other viverrids of the island it is not bound to the
tropical rainforests. It is not viewed as greatly at present but is definitely declining in numbers IUCN (International Union for the
Conservation of Nature and Natural Resources, 1978). Size of territories or hunting areas for C. ferox have not been determined, but in
dense populations, each animal is estimated to require 1 km². The tick, Haemaphysalis obtusa, is the only know parasite. The zoo at
Montpelltier, France, was the first to succeed in breeding, and handrearing fossas outside of Madagascar. Other than at this zoo, few
fossas are held in captivity.
All C. ferox caught and observed were solitary except for one instance in which several males were noted with a female in estrus.
Courtship behavior of males has not been described. Females ready for copulation take a typical lowered stance during which the
outer genital area in extruded 2 to 3 cm. Copulation usually last for more than 1h, but can last as much as 165 minutes, and take place
on the ground or on horizontal branches. The female mews during copulation. Pregnancy is recognizable only a few days before birth
by swelling of the mammae. Young suckle for about 5 minutes at a time and put their forefeet on the belly of the mother while she lies
on her side. Observations of some of the ontogenic behavior of fossas in captivity include the first scent-marking by males (from
glands in the genital region) at 7 months of age and by females (from glands in the anal region) after 9 months. Probably young part
from their mother at 15 to 20 months of age. Cryptoprocta ferox is active mainly at dawn and at night, with a peak of activity in the
hours before dawn. They also have been observed in the field during the day. In captivity, C. ferox can change to diurnal activity. The
mating season (during spring months of September and October in wild C. ferox shifted to April and May in specimens maintained in
the "Zoologischer Garten" in Hamburg (Vosseler, 1929). Intraspecific communication consists of relatively long-lived scent marks
made by both sexes, a few acoustical signals (threatening call, mewing by female during copulation, purring). and visual signals
(expression of the face and the whole body), but nothing is know about their exact messages. Fossas move in a more digitigrade
fashion on the ground and more plantigrade on branches. They climb and jump easily. The forefeet are used to capture and pin prey
to the ground. Agonistic behavior has been observed only during mating when aggression, including threatening calls and postures,
are evident. Males are fierce fighters; a female was observed to attack a male in the field and the opponents attempted to bite each
other. Fights usually are of short duration, after which the loser flee and the winner follows for a few meters.
The diploid number of chromosomes is 42, those of other Viverridae range from 24 to 52, whereas those of Felidae range from 36 to 38.
Rumpler (pers. comm.) studied the karyotype and found no information "which would allow us to classify Cryptoprocta closer to the
viverrids than to the cats."
The name Cryptoprocta refers to the anus (procta) being hidden (crypto-) by an anal pouch. The vernacular name also is spelled
"fosa." We thank C. Smeenk (Rijksmuseum van Natuurlijke Historie, Leiden) and M. Tranier (Museum National d'Historie Naturelle,
Paris) for the opportunity to study their specimens. Thanks go, as well, to M. moller and some other friends for elemanating the worst
linguistic faults.
Did you know that fossas are a bit larger in captivity than in the wild, its because they get a better diet and care. In the wild is because
like most animals some day they get shortage of food.
Did you know that a fossa when they eat chickens they may also eat the feathers
Did you know a fossa heart rate is 140 beats per minute.
And today, there are 55 fossa in captivity worldwide, 13 of them in a zoo in Duisburg, Germany. But biologists have yet to sort out some basic issues of fossa gender (see "Wild Thing," right), and the animal's behavior in the wild is even more of a mystery. In the United States, only the San Diego and the San Antonio zoos have fossa in their collections.