The Theory of Complex Accumulations
 (Back to Top.) ____________________ (Site Navigation Page.)
(Back to Top.) ____________________ (Site Navigation Page.)![]()
In this Section
2.4 Stepped Changes in Evolution
Return to
A New Model of Evolution Home Page![]()
2.0 The Theory of Complex Accumulations
No principle of evolution appears more self-evident than that individuals will enhance fitness by accumulating complexity. This is how evolution seems to work. An individual accumulates a slight improvement. This might make it a more complex creature, but if it enables it to procure more offspring than its rivals fitness goes up. (Individuals do not accumulate genetic complexity in single lifetimes. If among a group of individuals those born with more complex attributes are reproductively favored in the struggle for life, over time life will evolve more complex forms.)
This idea matches human expectations.
Moreover, if we define the increase of complexity as accumulation of genetic distance in the direction in which time flows, and treat fitness as a genealogy over many generations, fitness will increase in the direction of accumulation (the direction of time) virtually by definition. (We think of the history of life as the history of organisms becoming progressively fitter.)

Yet increasing complexity might also carry a fitness cost, although this depends on the definition.
This test makes fitness a retrogressive rather than an immediate measure, but that is the price of objectivity. We cannot observe a strong or fleet individual and assume that attributes aesthetically pleasing to humans are fit biologically. We must measure the results to be sure. We can only measure results among subsequent, rather than immediate generations.
If we can assess fitness objectively by measuring the amount of ancestor DNA distributed among descendents, the greater the number of generations we measured this over the better the result should be.
Yet, while viable traits can persist unaltered over thousands of generations, unique genomes, including the non-genetic and non-expressed DNA become quickly diffused as types change.
For example, suppose an ancestral fish acquired a small novelty that made it a better fish. Then that ancestor was very fit, because if the design is sound fish species can survive unchanged for huge periods. But suppose that for less well-adapted fish the ancestral seas were already saturated with fit designs. So, the only hope of procuring a lineage lay not in the sea, but pushing onto land (by becoming an amphibian). Amphibians did penetrate land so this was a fit phenotype move. But that did not make the ancestor genotype fit over many generations.
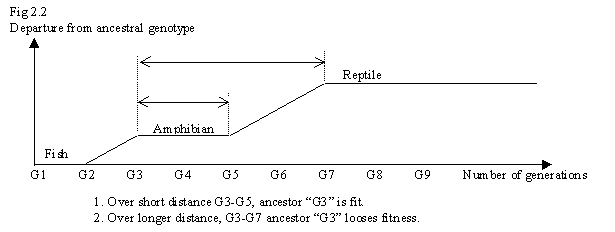
So, for each small step on the way from fish to reptile, for each fitness plateau of a stable intermediate species, fitness will rise. But over the entire ‘distance’ from fish to reptile, fitness will fall by a huge amount, which is what the new theory is saying. Fig 2.2 shows the problem. G3-G4, although only a transitory type, was stable enough that "G3" was a fit ancestor (to generations G4 and G5). But once major "vectoring" began again at G5, ancestor "G3" was no longer fit as an ancestral genotype.
Still, this is showing fitness falling as a relative measure. But does fitness also fall in absolute terms? In some ways, this question has no meaning, as fitness, ultimately, is a human measure of how we understand nature. But there is evidence that fitness does fall in absolute terms as complexity increases.
![]()
2.2 Absolute Fitness Falls
Fitness, always, is a human measure. In nature organisms do what they do, but modern humans, the intelligent creatures, care that what organisms do makes logical sense. One powerful method for humans to understand what organisms do is measure the amount of unique DNA than an organism, by its behavior, will pass into subsequent generations. Only this is not an easy measurement to take.
So most fitness modeling is instead done mathematically, and results compared to that observable among living species. These models show that if a modification to genotype improves adaptability, fitness will rise. Only such models can only be applied conclusively over a few generations, within the same species. But evolution has proceeded over billions of generations and millions of species. Over such huge distances, for what humans perceive as "improvements" (increase in complexity) fitness will fall. We can demonstrate (Fig 2.2) that fitness will fall as a relative measure, against an ancestor from a line that widely diverged from original type. But there is evidence that fitness falls absolutely for increase of complexity too.
There is also a theoretical argument that fitness must fall from early life, due to so-called "minimum complexity". There must be a minimum level of complexity for any DNA-coded cell to exist. Yet if there is a minimum state of complexity living reproduction can exist at, then any descendents at that state must be exact copies. There can be non-exact copies, but these would only add complexity to the minimal possible amount. We doubt if any primitive cell ever existed at minimum theoretical complexity. (A perfect state of anything is too unlikely in a heterogeneous universe!) Yet, the fittest organism ever was probably a cyanobacteria, which hardly changed its genotype in 3 billion years. But if this organism existed first, and had high fitness, and organisms that evolved later enjoyed less fitness, at some point organisms must have evolved against a loss of fitness.
Moreover, if fitness is passing of unique parent DNA into subsequent generations there are many reasons why fitness would fall as complexity increased. To suggest a few;
Only if fitness does fall as organisms become more complex, then we must find the reason. It is because in evolution there are both hard and easy to enact changes.
Explaining why is one of the most difficult to grasp of all modern concepts of evolution.
![]()
An early concept of evolution, partially adopted by Darwin, was that evolution was progress towards perfection, usually understood to be evolution of intelligent beings. Yet, modern studies show evolution is mostly non-progressive. Changes are more like oscillations than progress towards a goal. While ever organisms maintain or slightly improve fitness, this is the case. Again, it is because changes are always of two types.
Now why, in primal life, some bio-chemical properties are stable but hard to change and some properties are less stable but easy to vary, concerns the physics of the universe. But once life advances to DNA-coded multi-celled organisms the reason that some changes are difficult is statistical. Some changes will require large alterations to genome, many of which will be statistically unlikely to yield high fitness gains in a single generation. Only this argument is controversial. Opponents of evolution have argued that the eye, say, could not evolve because too many changes are required at once to produce the perfected organ. But while it contains hard-to-evolve attributes (muscles, nerves, photoreceptors) the eye assemblage is not itself a complex accumulation. It has been shown (many times) that each incremental improvement in vision, even for a partially perfected eye, always yields a slight fitness advantage. Plus eyes are not monophyletic. Several types of eye have evolved several different times, so even if we had no clue how eyes evolved, it must still be by a simple fitness path, because eyes evolved so many times.
But take a genuinely monophyletic attribute, such as the four-chambered heart.
Although it sounds involved, growing the tissue dividing the chambers is not itself complex, as even a part divided chamber will give a more efficient blood flow than no divide at all. Yet despite that it appears a simple adaptation to fully divide the ventricle, no creature other than a mammal has ever done so, or ever will. The reason is that while a fully divided ventricle will yield more efficient blood flow, it will not yield an overall fitness advantage without extensive modification to lungs, skin, arteries, breathing, and endothermic regulation. This means that he four-chamber heart comes not as an isolated improvement, but part of a repertoire of modifications to a large number of attributes, each of which must work with the other to yield a high fitness gain. Only each attribute has its own slight improvement, and in competition with a neighbor, even the slightest improvements are passed on. So, although we can see why these attributes would also evolve in isolation, we must explain why they evolved only once.
Again, it has to do with fitness, and that some changes are easy, but others are hard to evolve. Take the case of reptilian predators, also in competition with each other, for a very slight advantage. Now extending the length of anything is an easy-to-enact change. In a single generation, one reptile might produce a slightly longer claw, the other a slightly extended ventricle divide. Both improvements can be produced, but under the prevailing conditions of competition, which is more likely to be fit?
The four-chamber heart is a complex accumulation them, but not because it is complex to extend the ventricle divide. Rather, extending this alone yields low fitness in isolation from other changes, compared to self-supporting "fitness rich" adaptations an individual can undergo. Yet all attributes that are monophyletic (evolved at one source) and homologous (radiated from the source into many types) are complex accumulations. Each attribute has a small fitness gain. But the big gains are long term, and involve modifying many features, which requires evolution over a huge genetic distance. But as we also saw, large changes of genome eventually result in fitness loss, as a relative measure for ancestors, but fitness also falls absolutely for modern types, for many reasons. To these reasons we can now add two more.
Note: The concern over fitness loss among ancestors is not over the "feelings" of the ancestor, who has long perished. A "retrogressive" fitness loss means that a large portion of the ancestor's genes turned out to be not suitable for modern conditions. This forces the evolutionary cost of further adaptation onto descendents.
![]()
2.4 Stepped Changes in Evolution
Contention over whether evolution proceeds by continuous small steps or consists of long periods of stasis "punctuated" by sudden increases in species complexity is now well debated. However, most issues concern interpretation of the fossil record and relative rates of evolution. Yet, no matter which view one holds, sudden, large mutations at many alleles at once are statistically unlikely to be fit in a single generation. So, if evolution does proceed in a stepped manner, then small changes accumulating over long periods must somehow be causing the stepped effect.
We can show stasis and the "stepped" changes as an effect of fitness, again to do with all changes in evolution being either easy or hard to evolve.
For example, in hundreds of millions of years no descendent of a shark could afford the fitness penalty of evolving into anything other than a more efficient shark. The ancestral shark design was already very fit, and sea conditions are mostly stable, so fitness among quick, easy-to-enact, single generation improvements, retains a shark always to be a shark.
Yet while it never happened to sharks, many events can temporarily "loosen" the strict fitness bind that sharks say, are always in. In stable conditions, not only a single species (sharks) but all the species within an environmental range will be in fierce competition for very slight advantage. This creates a range "equilibrium" in which each individual in each species in the range can only afford easy-to-enact fitness enhancements to stay ahead. (It is not certain if this is what Steven Gould meant by equilibrium. It is never this sharp, but easy and hard to enact changes form an on-off, either-or, "state". So species within a range being all restricted to easy-to-enact changes is a "state" of equilibrium, defined perhaps with a sharpness of distinction no biologist could accept.)
However, the equilibrium of any range can be "punctuated" by unsettling events, like environmental or geological change, vacation of ranges by extinction of rival species, invasion of ranges, overpopulation, migration to new ranges, and so on. When this occurs the fitness rules become loosened.
Loosening of fitness rule favors complex accumulations, because these consist of many small, low fitness yield changes. But although many changes are required they will tend to be in spurts. While genotypes are changing and fitness yields are low (yields are low because types are changing) the species will be "searching" for a new equilibrium plateau, where high gain, easy-to-enact yields will again predominate. (Roughly, the founder individual who adapts to the new plateau best will enjoy high fitness. But until the new plateau is found, no ancestor will enjoy high fitness because every few generations the founder type must alter again, until the process stabilizes.)
Stepped changes in evolution are not hard to explain, once we accept that all changes are either easy or hard to enact changes, and that fitness falls as species become more complex.
But if fitness yields for changes are low, and conditions have not stabilized, there will be much experimentation and many changes until a new stability is found. This will appear in the fossil record as a "stepped" or relatively rapid rate of change, although there are other ways to explain this. (Again, this is phylogenic explanation of the stepped effect. There can be random changes of climate or geology, or migration and extinction for non-phylogenic reasons.)
![]()