The 'Two-Axis' Model of Evolution
No PDF version of this file, which is large because of diagrams, but it can be provided on request.
(Note: This essay is philosophical, more on how humans model nature than how evolution works. See other essays in
new model of evolution for details of evolutionary processes leading to the 'two-axis' model.) (Back to Top.) ____________________ (Site Navigation Page.)
(Back to Top.) ____________________ (Site Navigation Page.)![]()
In this Section:
6.1 Modeling Nature All models of nature contain fixed and variable traits.
6.2 Fixed and Variable Processes Fixed traits evolve from variable ones once only!
6.3 Phase Transitions Evolution of fixed traits is a phase transition.
6.4 Evolution along Two Axes We can show the effects best on a 'two-axis' model.
Return to
A New Model of Evolution Home Page![]()
6.0 The 'Two-Axis' Model of Evolution
Models that humans make of nature are never exactly how nature works, in all its detail and nuances.
So, although nature is a single process, humans never use a single model of it. Rather, we must choose among available models one most apt to that segment of nature that we are trying to explain. Yet, although we have different models of nature, all models share a universal characteristic. This is that the models compose of both;
We can show this universal characteristic of some traits always varying against other traits that are fixed, on a diagram (Fig 1.1). Variable properties alter left-to-right along an axis that we associate with the flow of time. The "fixed", non-variable properties, we show "rotated" 900 away from the plane in which time flows. The diagram is as follows: (For "trait" read property, attribute, etc.)
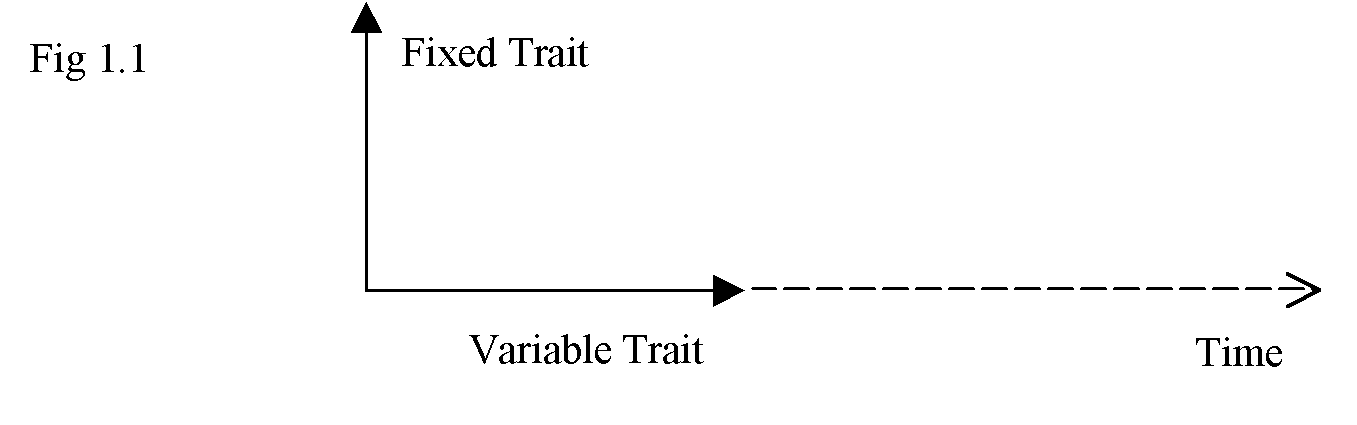
Variable properties are those we measure against properties we regard as fixed (we navigate against the "fixed" stars). This applies to models of evolution too;
Yet often overlooked (and for thousands of years was not known) is that since the beginning of the universe properties now fixed have also evolved, from a state of being variable. For example;
So, apart from fundamental constants (such as the speed of light or Plank's Constant) most properties in life and the universe we now regard as fixed were once evolving. This makes evolution of complex entities such as stars or planetary systems, but especially the evolution of life, a dual process.
So, any universal model of evolution for any point in time cannot cover just evolution of the foreground properties. It must include the background processes of evolution too, in which properties once variable themselves evolve into the modern fixed properties of nature.
6.2 Fixed and Variable Processes
Every process of nature involves variable "foreground" properties altering against fixed "background" ones. This would be the case even in instances when the fixed "background" properties themselves evolved the first time. (There would be a set of even deeper 'background" properties, like a stable atomic structure.) Only strangely, whereas the model is still the same where "background" properties evolve, it is different in that "background" evolution only occurs once!
Yet, for "foreground" evolution the same cycle of change occurs many times. Matter only evolved once, but different assemblages of matter are recycled trillions of times over. Or while the DNA code only evolved once, the code sequence alters every gamete in uncountable trillions of combinations.
Another controversial difference is the teleology of the processes. Although we call the "foreground" process evolution, most changes are cyclic or oscillatory. Like the seasons, or birth and death, changes occur. But once the cycle is complete (the life cycle of a star or a species) we cannot measure the change as progress towards a goal. This often called the Red Queen Effect, because "you have to run as fast as you can just to stay in the one place". Yet, while foreground evolution does not result in scientifically measurable progress, background evolution is progressive in a historical sense.
Again, we can show the effect on a simple diagram. At any point in time some properties change against a background set of properties, not varying at the same rate. But as the universe changes over time, certain properties do change in unique ways. This is diagrammatic, but maturing properties appear as a 'rotation' through 900, from the axis of variability to axis of fixation. Fig 1.2 shows the effect.
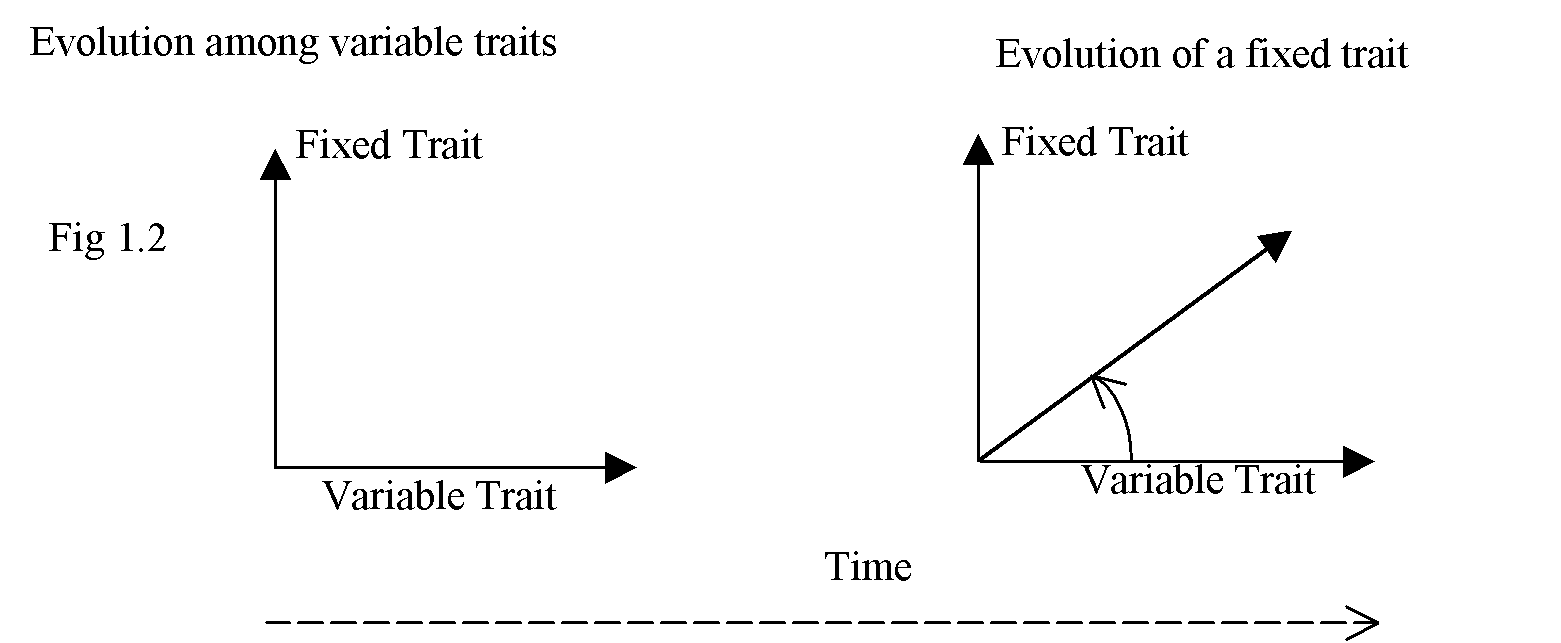
Even so, this diagrammatic 'rotation' only involves some elements.
Plus most cataclysmic changes in evolution on Earth were in the remote past. So, by the time intelligent beings evolve only "foreground" evolution remains for them to study as living evolution. (The fossil record supports a theory of large-scale, cataclysmic changes. But living evolution is of small-scale changes.) So, we dispute whether changes in the past were "foreground' evolution over longer times, or if different processes operated in the past than today.
But the issue is complex. Whether stars are Type I or II, the laws of physics and chemistry are the same. And whether changes in evolution are small or large, laws of natural selection or population genetics do not change either. But;
Similarly, the physical laws of the universe are invariant with time. But which processes are possible in the universe depend on its size, density, and matter-energy composition, which alter over time.
So while the laws of physics, or laws of life, do not appear to alter over time the conditions in which these laws act do alter and have a history, which is difficult to model.
![]()
One noticeable effect of human evolution is a major change from evolution by adapting biology, to evolution by adapting culture and behavior, while leaving the biology to change imperceptibly. To reach this point, human biological evolution was very rapid, but this makes the change even more distinctive once it occurs. In the physical sciences, we would call such a transition from one state of alteration, to alteration under a different state, a "phase" transition, or phase shift. We suspect that such a phase transition occurred in human evolution, but we have no idea of;
The new model can solve problem b) by demonstrating that phase transitions, or phase shifts (we will use the latter term) occur throughout evolution. Human evolution was just the last such transition, in a series since life began. For living matter, a phase shift is the state of a property changing in early evolution, but stabilizing in a later epoch. For example;
Human evolution then was not a "phase shift" in that biology was transmuted into culture, although that was the effect. It was a phase shift biologically, in that an intelligent, culturally adaptable creature had to first evolve. But once it evolved its type matured, and was able to radiate into a huge number of adaptive (mainly cultural) forms of existence.
More difficult is problem a), concerning which physical property is involved in transitions. Until we measure a property changing, the term "phase shift" is ill defined. The DNA sequence expressing the H4 protein has been stable for at least 1012 generations. But the sequence had to evolve, so if it once exhibited a stability of less than 1012 and then went through a transition to the new stability this was a measurable phase shift. (H4 evolved in the remote past, so we do not know when the shift occurred.) Yet many attributes have evolved in shorter evolutionary times than the subsequent stability they produced. For example;
Even so, any alteration to rates of mutation in DNA is only measuring an effect. We could use a "selfish gene" argument that phase shifts of mutation rates from an evolving to a stable mode was a strategy of genes, to retain high sequence conservation throughout subsequent evolution. (This is the "smart gene" hypothesis, earlier.) But we could not explain all shifts in mutation rates as caused by gene selfishness.
Also high sequence conservation seems more a result of selection than inherent phase shifts, because non-expressed (hence non-selected) sequences are always less conserved than expressed ones, and so they show less of a "phase shift" effect. (See also the section of phase shifts in the "smart gene" hypothesis.)
So, for our present understanding a phase shift is an event rather than a cause. (Expansion of the universe is a cause, while a phase transition from energy into matter is an event resulting from the cause.) For biological evolution a "phase shift" is an event in the history of life in which a trait once evolving matures and stabilizes.
No matter how far back we find everywhere traits that evolved rapidly in a previous age stabilizing as mature traits for the next stage of evolution. Only for living tissue there is no property that can consistently quantify a difference between fixation and variability.
Still, we can quantify mutation rate as an angle.
We can interpret this 'angle' to mean that while a gene is within the "average" mutation rate (10-6) it is partially variable and a partially fixed. Allele frequencies will be changing every generation along the 00 horizontal axis, which is the axis of variable traits. At 450 to horizontal, mutation in a gene might effect the expressed traits of a species. But once the mutation rate slows to 10-7 or lower, it moves outside of "residence" of that species. It will mutate too slowly to appear in a small population, within a random window of selective pressure to change. Fig 5.2 (from "smart gene" hypothesis) shows the effect.

Critics have objected that this 'angle' argument is false; the tan-1 formula is a contrivance to "force" a bipolar effect, when none is there. Yet, the effect exists because some properties are variable while background properties are "fixed".
But genes did not physically rotate. It is just a representation. (In an alien civilization, time does not flow left to right across a page, positive rotation might not be counter-clockwise, and angles will not be measured in degrees.) But no matter how we present it, throughout the universe properties now fixed once evolved. We need to represent this effect with a model appropriate to our understanding.
Only in evolution traits group into easy and hard to evolve types. We have examined this, and found that;
This infers that "rotation" from a shallow to a steep angle would be an irreversible event. That is, the rotation would only ever be counter-clockwise. This is different from cyclic processes in evolution, like increase of decrease of length, or point mutation of a base, which are reversible. In this sense, the processes of adapting easy-to-alter and evolving hard-to-alter traits in evolution are distinct, and the 'two-axis' model is only a representation of that distinction.
6.4 Evolution along Two Axes
If one wished to describe in nature two processes, one the inverse of another, our mathematics and language too readily provide the tools and terms to use. Language, especially Greek and Latin roots use many inverse terms.
Mathematics too is replete with inverse terms.
Such coincidences common throughout science create grave debate. We wonder if the universe is naturally bi-symmetric, and our language and mathematics reflects that, or if our mathematics and language is contrived by human prejudice and through its prism we impose a "world view" on nature.
Another problem is limited human observation. We have only seen evolution on one planet. Even then, we discovered evolution almost immediately after intelligent life (us) emerged, so the story of evolution appears to end with its own discovery.
But then what happens?
We cannot answer this because no human has witnessed it. Perhaps
In these cases, all attempts (and there are many, not just this theory) to derive the "formula" of why intelligent life evolved are as vain as attempts to derive why God specially created it.
This is why if we think there is a difference, we must demonstrate that "foreground" evolution is analytically different from the "background" process objectively, independent of human perceptions, language, or use of terms. Yet in the universe, objectively, some properties are easier to alter than others. For example;
Only once the DNA code evolves, human perceptions again distort this bi-symmetric model of evolution. Humans have difficulty understanding why, if any DNA-coded trait is easy to evolve, DNA-coded traits will continue to group as easy or hard to evolve. We can only point out that;
This gives fitness an additional role. We think of fitness and selection refining or improving design. But fitness and selection also retain that feature of all existence that some properties of the universe are easier to change than others. Evolution is a more efficient process overall, if it proceeds this way.
But invoking fitness to explain how easy-to-enact changes are favored strikes another conceptual difficulty explaining how the hard-to-form traits evolve.
This last effect is very strange. There is evidence that as life becomes more complex fitness falls. It does for sex, and it appears to fall for evolution of eukaryote from prokaryote life. But it is harder to "see" how for each major increase in complexity (as for evolution of a reptile into a mammal) fitness will fall. Mathematically, this requires that a series of small positive fitness gains "add up" to an overall fitness loss. This should not be a problem, because such effects are frequent in the physical sciences, but it confounds conventional notions of evolution.
This is why we need the 'two-axis' model.
Postulation of a 'two-axis' model for evolution does not "cause" fitness to fall as a lineage of types accumulates complexity. But fitness falls in nature, and the 'two-axis' model helps explain why. In Fig 6.3 we see two views of how small changes accumulate. In the first, we assume all changes are along the 00 'axis'. And all small changes first accumulate this way, as events in time. But as new traits, novelties or sequences mature and solidify, they "rotate" 900 away from the axis of time, to indicate that they are not subject to further change. This is acknowledged anyway, as most changes are "foreground" against a background of traits that are fixed. So, the diagram shows that the individual is a composite of homoplastic traits, and deeper homologous traits, and the traits alter on different time scales. Already we see that small foreground changes will not add linearly.
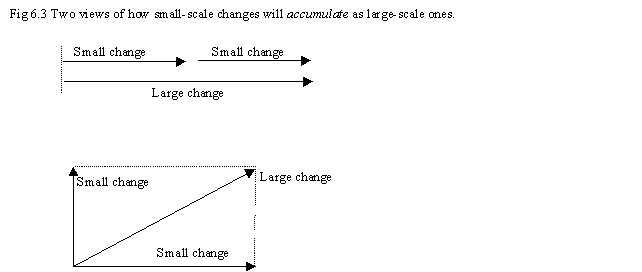
Yet as life evolves greater complexity, more attributes "phase shift" to the vertical axis. The huge diversity of mammals is expressed from a core set of 70% of genes. And Great Ape diversity, and fast evolution into different types, is expressed from a core set of 96% of genes. So as life becomes more complex, different types can evolve, faster, from a smaller core set of genes and fixed attributes. (This data needs checking! Mammals are complex, and they can evolve quickly into a huge range of diverse types, while retaining a core set of genes and traits in common. It needs to be checked how fish, reptiles or other classes compare. Possibly once more data is known, this thesis might need updating.)
Fig 6.4 shows the problem. Complex life contains a huge genome, but most of it is unexpressed or non-coding DNA, which is tied to individuals, but is not very fit as exact copy. On the other hand the useful, core traits of complex life are expressed by a tiny fraction of DNA, most of it sharing a high homology among a variety of types. (The composite diagram would be more pronounced. For a 10-5 "average" mutation rate genes mutating slower would be near vertical. Non-coded DNA might be altering once per generation for sexual reproduction, so it would be drawn horizontal. But different scales would show the effect differently.)
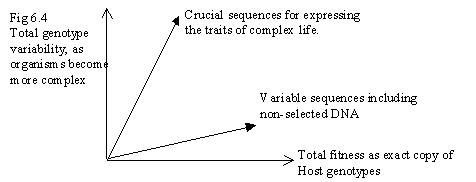
So, there are not two separate processes in evolution, and life is not evolving along two 'axes'.
Humans have to explain this too. They have to explain why a property that always adds as a positive (fitness) can result in a negative as a summation, and explain why a property that is infinitely variable in isolation, can saturate in summation.
The 'two-axis' model of evolution is an attempt to represent, diagrammatically, how these broader, more complex processes of evolution work. (See also notes on Complex Time)
![]()